Dr. David Eisner discussed mechanisms of calcium regulation in the heart, focusing on the calcium regulating ryanodine receptor (RyR) and its relationship to cardiac function and cardiac arrhythmias. Electrical stimulation of cardiac myocytes causes a transient increase in calcium, which activates myocyte contraction. Myocyte calcium levels are low (about 100 nM) when at rest during diastole and increase to a few hundred nM during systole. The major factor that regulates the force of the contraction is the size or amplitude of the systolic calcium transient.
|
Figure 1. Calcium Induced Calcium Release from SR
【Click to enlarge】 |
|
|
One source of the calcium transient is the surface membrane L-type calcium current. However, most of the calcium that activates contraction comes from the sarcoplasmic reticulum (SR). Calcium is released from the SR into the cytoplasm by the specialized release channel, RyR. The probability that the RyR is open (Po) is increased by increased cytoplasmic calcium. The action potential activates calcium entry into the cell, some of which binds to the RyR causing it to open and release more calcium into the cytoplasm (Figure 1). This process is known as calcium-induced calcium release. The increased calcium activates the myofilaments, producing a contraction. The myofilaments relax when cytoplasmic calcium is reduced. Calcium is pumped back into the SR by SR Ca-ATPase (SERCA), which is regulated by cytoplasmic calcium and the accessory protein phospholamban. Phosphorylation of phospholamban, as occurs during beta adrenergic stimulation, increases SERCA activity. Calcium also is pumped out of the cytoplasm by the sodium-calcium exchange (NCX), which generates the electric current. On each heartbeat, the influx of calcium into the cell equals the efflux. Thus, in the steady state, sarcolemmal calcium entry equals calcium exit.
The size of the calcium transient that enters the cytoplasm is regulated by the amount of calcium in the SR, RyR properties, and the amount of calcium entering by the calcium current. The amount of calcium in the SR is the major regulator of the calcium transient size. The size of the calcium transient is proportional to the third power of SR calcium content (SR Ca)3. Even a small change in SR calcium content has a large effect on the size of the calcium transient.
The major factor controlling SR calcium content is autoregulation—the release of calcium from the SR affects movement of calcium across the sarcolemma, which in turn affects the amount of calcium in the SR. When the SR is emptied of calcium, the L-type current inactivates more slowly than when there is calcium in the SR. The bigger the calcium transient in the SR, the less calcium enters on depolarization. The amount of calcium pumped by the NCX also depends on the size of the systolic calcium transient. The bigger the calcium transient, the more calcium is pumped out of the cell.
The RyR Po is increased by phosphorylation, cADP ribose, caffeine, and free radicals, and decreased by acidosis, tetracaine, and heart failure. Beta adrenergic stimulation by catecholamines increases the force of contraction. A recent hypothesis suggested that beta adrenergic stimulation increases systolic calcium by phosphorylating the RyR. In an experiment to test this hypothesis, low concentrations of caffeine were applied to phosphorylate the RyR. As a result, more calcium entered the SR. Calcium influx remained constant, but because the calcium transients were bigger, the NCX was more activated and calcium efflux increased. With the calcium efflux greater than the influx, the SR calcium content continued to decrease until steady state was reached. This experiment shows that altering RyR properties in the steady state has no effect on the size of the calcium transient. Dr. Eisner concluded that it is unlikely that RyR phosphorylation is involved in the positive inotropic effects of beta adrenergic stimulation.
Heart failure has been treated with positive inotropes to increase SR and systolic calcium. However, positive inotropic agents that increase calcium induce more arrhythmias, a common cause of death in patients with heart failure. When the SR is calcium overloaded, calcium is released from one part of the SR and propagates as a wave of calcium-induced calcium release. Such calcium waves are arrhythmogenic. The wave occurs in diastole, resulting in calcium efflux on the NCX, which produces a depolarizing current during diastole. The consequence of this is that calcium waves produce extrasystoles by a delayed after depolarization (DAD) mechanism.
Arrhythmias due to SR calcium overload are known to be caused by digitalis toxicity, reperfusion, and inotropic agents. Recent evidence has shown that these arrhythmias also occur if RyR properties are altered so that the RyR leaks calcium and the threshold is decreased. This situation occurs in catecholaminergic polymorphic ventricular tachycardia (CPVT) caused by RyR mutations and in heart failure patients when the RyR is phosphorylated.
|
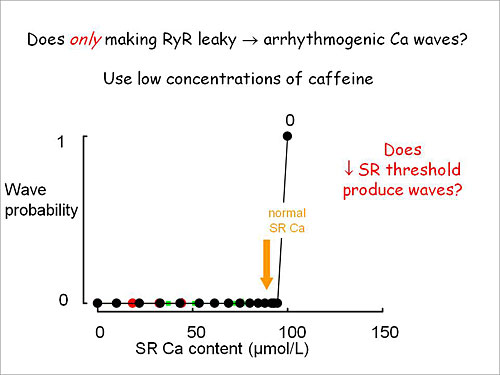
Figure 2. Does only making RyR leaky cause arrhythmogenic Ca waves?
【Click to enlarge】 |
|
|
Dr. Eisner showed that adding caffeine to cardiac myocytes decreases the threshold of wave probability as a function of SR calcium content (Figure 2). Measures of cytoplasmic calcium showed that when a low concentration of caffeine was added, waves initially appeared but in the steady state there were no diastolic waves. Initially the calcium measurements were steady but the caffeine-induced waves caused calcium efflux from the cell, decreasing the SR calcium content and the waves disappeared. When the same experiment was done in the presence of isoproterenol, addition of caffeine induced calcium waves that persisted in the steady state.
|
Figure 3. The presence of waves depends on the interplay between SR content and SR threshold for waves
【Click to enlarge】 |
|
|
Further experiments showed that beta-adrenergic stimulation increases the SR calcium, primarily by increasing SERCA activity. There appears to be a threshold for waves. In the absence of isoproterenol, the SR calcium content is below this threshold; in the presence of isoproterenol, SERCA is stimulated and SR calcium increases. Dr. Eisner concluded that beta-adrenergic stimulation acts by increasing SR calcium content, not by decreasing the threshold. Waves only appear with the presence of both caffeine (which lowers the threshold) and isoproterenol (which increases SR calcium content) (Figure 3). This appears to be the same mechanism that produces arrhythmias in children with CPVT, in which mutant RyR lowers the SR threshold and stress increases SR calcium.
Beta blocker therapy is effective in about 60% of children with CPVT. The investigational agent JTV519 restores RyR function and blocks arrhythmias but has no effect in a mouse RyR model of CPVT. Dr. Eisner found that depressing the RyR opening with tetracaine eliminates arrhythmogenic diastolic calcium release caused by isoproterenol but increases systolic release. While tetracaine is not selective and blocks all sodium channels in the body, these experiments pave the way for development of an agent that is much more selective for the RyR.
|